Yessica Llamas González*
Introducción
La Organización Mundial de la Salud considera que varias enfermedades tropicales causadas por arbovirus están “desatendidas” y siguen siendo un desafío para la salud pública. Las infecciones por arbovirus se propagan a nivel mundial como resultado de la expansión de los mosquitos que, a su vez, esto es causado por el aumento del comercio y el cambio climático [1].
Los alfavirus son miembros de la familia Togaviridae que se transmiten principalmente por la picadura de artrópodos hematófagos. Clásicamente, los alfavirus se dividen en alfavirus del Nuevo Mundo y del Viejo Mundo según su abundancia geográfica histórica [2]. Los alfavirus del Nuevo Mundo causan enfermedades encefalíticas e incluyen los virus de la Encefalitis Equina Venezolana y Occidental (VEEV y WEEV) y el virus Mayaro (MYV). Las infecciones por alfavirus del Viejo Mundo, como el chikungunya (CHIKV) y el virus Ross River (RRV), resultan principalmente en una enfermedad reumática que causa un dolor debilitante en las articulaciones.
MAYV es un alfavirus artritógenico reemergente del Nuevo Mundo [3] (L-Smith, 2018). Este virus se aisló por primera vez de muestras de suero de trabajadores forestales en Trinidad en 1954 [4]. MAYV es un virus que puede infectar, replicarse y diseminarse tanto en hospedadores vertebrados como en invertebrados. En los seres humanos, el virus causa la fiebre de Mayaro, que se caracteriza por una artralgia de larga duración similar a la que ocurre en la fiebre del chikungunya.
La fiebre de Mayaro puede convertirse en un importante problema de salud pública, particularmente en las áreas rurales, con una prevalencia creciente en la región amazónica debido a cambios en los ecosistemas. MAYV ahora se está extendiendo rápidamente a otras regiones de América Latina. Es probable que la carga mundial de la fiebre de Mayaro todavía se subestime en gran medida debido a la falta de diagnósticos adecuados y precisos. Al mismo tiempo, el mosquito Aedes, que puede propagar MAYV y otros virus, está invadiendo nuevos hábitats y regiones a un ritmo alarmante.
Manifestaciones Clínicas
Filogenéticamente, MAYV está estrechamente relacionado con CHIKV y, como este último, causa una enfermedad debilitante, similar a la gripe en el hospedero infectado, que es indistinguible de la fiebre Chikungunya. Los síntomas principales incluyen escalofríos, fiebre, manifestaciones gastrointestinales, dolor ocular, mialgia y artralgia. En particular, la artralgia puede durar meses o años, lo que hace que la fiebre de Mayaro sea aún más debilitante que las infecciones por flavivirus, cuyos síntomas duran solo de una a tres semanas [5]. El mareo y la picazón son las otras manifestaciones clínicas de la enfermedad. Pueden producirse complicaciones graves debido a la infección por MAYV, entre las que se encuentran miocarditis, manifestaciones hemorrágicas y neurológicas. Como ocurre con otros arbovirus, no se dispone de un tratamiento específico para la infección por MAYV y solo se ofrece atención de apoyo a los pacientes, con el objetivo de tratar el dolor y la fiebre con analgésicos y/o antiinflamatorios no esteroideos (AINE) [6].
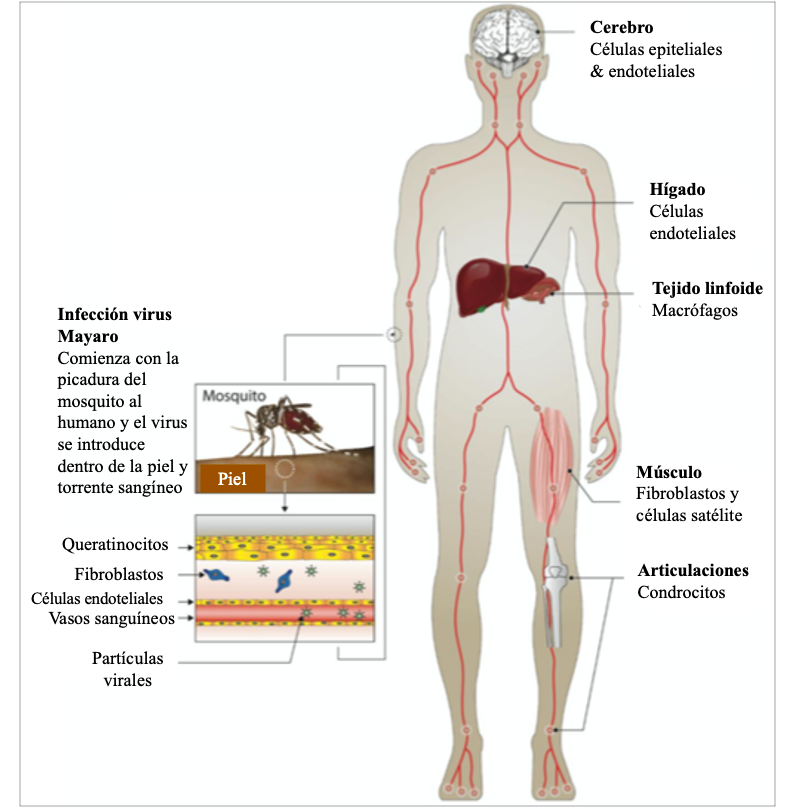
Figura 1. Diagrama que muestra la probable diseminación del MAYV en el ser humano. La transmisión del MAYV se produce tras su inoculación por un mosquito infectado (Haemagogus janthinomys, Aedes aegypti, etc.). A continuación, el virus se replica en la piel (más concretamente en el lugar de inoculación del virus por el vector competente) y se propaga a los tejidos diana (músculo, hígado, articulaciones, etc.) a través de los numerosos vasos sanguíneos, seguido del reclutamiento de células inflamatorias en estos tejidos.
Estructura Molecular
MAYV tiene un genoma de RNA monocatenario de sentido positivo de 11.5 kb con dos marcos de lectura abiertos que codifican las proteínas no estructurales (nsP1, nsP2, nsP3 y nsP4), que están implicadas en la transcripción y replicación del RNA viral y en la estructura de proteínas (C,E3, E2, 6K y E1), algunas de las cuales forman parte de la partícula viral [7].
Mecanismos de Evasión Viral
Los virus son patógenos intracelulares obligados que han desarrollado diferentes estrategias para replicarse en las células huésped, incluido el secuestro de la vía ubiquitina-proteasoma (UPS). La ubiquitinación consiste en la conjugación de moléulas de ubiquitina con un residuo de lisina en una proteína sustrato [8]. El UPS regula diferentes procesos celulares, como la degradación de proteínas, la reparación del DNA, el tráfico de receptores, la progresión del ciclo celular, la inmunidad y la autofagia [9,10]. No es sorprendente que diferentes virus como el virus de la hepatitis E (HEV), el rotavirus (RotV), el citomegalovirus humano (HCMV), el circovirus porcino tipo 2 (PCV2), el virus de la influenza A (IAV) y el virus respiratorio humano virus sincitial (HRSV) explotan el UPS para favorecer su replicación [11]. Además, varias líneas de evidencia indican que los alfavirus manipulan el UPS para promover su replicación. Se ha observado que las proteínas nsP2 del virus Sindbis de los alfavirus del Viejo Mundo (SINV) [12], el virus Selimki Forest (SFV) y CHIKV utilizan un mecanismo dependiente del proteasoma para provocar la degradación de Rpb1, una subunidad catalítica de la RNA polimeras II que induce la inhibición de la transcripción celular [13,14,15]. En estudios recientes se ha reportado que la proteína de la cápside de VEEV está ubiquitinada y que esta modificación es importante para su replicación. Sin embargo, aún no se ha explorado el papel del UPS en la replicación de MAYV. El objetivo de este estudio fue evaluar el papel del UPS en la replicación de MAYV con la implementación de moléculas inhibidoras como MG132 y Lactacistina, que podrían servir como futura terapia para el tratamiento de la fiebre Mayaro.
En este estudio se evaluó el efecto de los inhibidores del proteasoma sobre la replicación de MAYV. Se llevó a cabo una tinción por inmunofluorescencia de células HeLa infectadas con MAYV que fueron tratadas o no con los inhibidores del proteasoma usando anticuerpos anti-MAYV. Se observó una reducción significativa en el porcentaje de células positivas para MAYV después del tratamiento con los inhibidores del proteasoma como se muestra en la Figura 2. Estos datos indican que la inhibición del proteasoma reduce la carga de la progenie de MAYV.
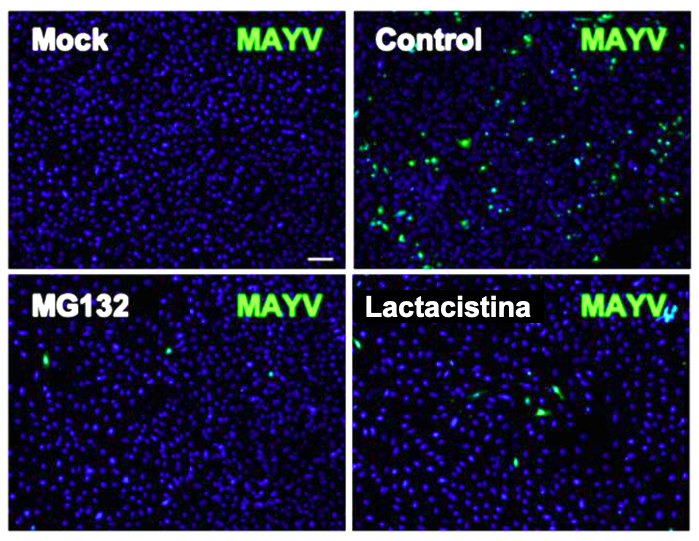
Figura 2. Células HeLa tratadas o no con los inhibidores del proteasoma que se infectaron con MAYV. Las imágenes se obtuvieron con un microscopio confocal y se analizaron con el software ImageJ.
Conclusión
Los virus emplean la maquinaria y los recursos de la célula huésped para facilitar su replicación. Las vías de degradación proteasomal desempeñan un papel fundamental en la regulación de múltiples funciones de proteínas y son esenciales para muchos procesos celulares, incluida la progresión del ciclo celular, la inmunidad y la autofagia. Por lo tanto, los virus han evolucionado para secuestrar la maquinaria proteasomal del huésped para mejorar su replicación.
MAYV es un alfavirus considerado como un patógeno emergente en América Latina que aún se desconocen los factores esenciales del huésped involucrados en su replicación. En este estudio se investigó el papel del UPS en la replicación de dicho virus desatendido. Los resultados indican que la inhibición del proteasoma reduce la producción de partículas virales infecciosas de MAYV. Los inhibidores del proteasoma MG132 y Lactacistina podrían ser un blanco farmacológico para controlar la infección por MAYV.
*Yessica Llamas González. Químico Farmacobiólogo de profesión, egresada de la Universidad de Guadalajara. Completó sus estudios de posgrado en la Universidad de la República de Uruguay; actualmente es candidata a recibir el grado de Doctor en Ciencias Biológicas, con especialidad en Virología. Participación en congresos internacionales en Panamá y Barcelona.
Referencias
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antiviral Res. 2010, 85, 328–345.
- Rana Abdelnabi and Leen Delang. Antiviral Strategies againts Arthritogenic Alphaviruses. Microorganisms. 2020, 8, 1365. Doi:10.3390/microorganisms8091365.
- Jessica L. Smith, Christine L. Pugh, Emily D. Cisney, Sarah L. Keasey, Carolina Guevara, Julia S. Ampuero, Guillermo Comach, Doris Gomez, Margarita Ochoa-Diaz, Robert D. Hontz, Robert G. Ulrich. Human Antibody Responses to Emerging Mayaro Virus and Cocirculating Alphavirus Infections Examined by Using Structural Proteins from Nine New and Old World Lineages. Clinical Science and Epidemiology. 2018. 3, 1-14.
- Anderson C.R., Downs W.G., Wattley G.H., Ahin N.W., Reese A.A. Mayaro virus: a new human disease agent. II. Isolation from blood of patients in Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1957;6:1012–1016. doi: 10.4269/ajtmh. 1957.6.1012.
- Levi, L.I.; Vignuzzi, M. Arthritogenic Alphaviruses: A Worldwide Emerging Threat? Microorganisms 2019, 7, 133.
- Aguilar-Luis, M.A.; del Valle-Mendoza, J.; Silva-Caso, W.; Gil-Ramirez, T.; Levy-Blitchtein, S.; Bazán-Mayra, J.; Zavaleta-Gavidia, V.; Cornejo-Pacherres, D.; Palomares-Reyes, C.; del Valle, L.J. An emerging public health threat: Mayaro virus increases its distribution in Peru. Int. J. Infect. Dis. 2020, 92, 253–258.
- Acosta-Ampudia Y., Monsalve D.M., Rodriguez Y., Pacheco Y., Anaya J.-M., Ramirez-Santana C. Mayaro: an emerging viral threat? Emerg. Microbes Infect. 2018;7:163. doi: 10.1038/s41426-018-0163-5.
- Clague M.J., Urbé S. Ubiquitin: Same molecule, different degradation pathways. Cell. 2010;143:682–685. doi: 10.1016/j.cell.2010.11.012.
- Swatek K.N., Komander D. Ubiquitin modifications. Cell Res. 2016;26:399–422. doi: 10.1038/cr.2016.39
- Viswanathan K., Früh K., DeFilippis V. Viral hijacking of the host ubiquitin system to evade interferon responses. Curr. Opin. Microbiol. 2010;13:517–523. doi: 10.1016/j.mib.2010.05.012.
- Barrado-Gil L., Galindo I., Martínez-Alonso D., Viedma S., Alonso C. The ubiquitin-proteasome system is required for African swine fever replication. PLoS ONE. 2017;12:e0189741. doi: 10.1371/journal.pone.0189741.
- Akhrymuk I., Kulemzin S.V., Frolova E.I. Evasion of the Innate Immune Response: the Old World Alphavirus nsP2 Protein Induces Rapid Degradation of Rpb1, a Catalytic Subunit of RNA Polymerase II. J. Virol. 2012;86:7180–7191. doi: 10.1128/JVI.00541-12.
- Cheng S., Yan W., Gu W., He Q. The ubiquitin-proteasome system is required for the early stages of porcine circovirus type 2 replication. Virology. 2014;456–457:198–204. doi: 10.1016/j.virol.2014.03.028.
- Widjaja I., de Vries E., Tscherne D.M., Garcia-Sastre A., Rottier P.J.M., de Haan C.A.M. Inhibition of the Ubiquitin-Proteasome System Affects Influenza A Virus Infection at a Postfusion Step. J. Virol. 2010;84:9625–9631. doi: 10.1128/JVI.01048-10.
- Lupfer C., Pastey M.K. Decreased replication of human respiratory syncytial virus treated with the proteasome inhibitor MG-132. Virus Res. 2010;149:36–41. doi: 10.1016/j.virusres.2009.12.010.